Life History & Behaviour
Most of the mechanisms used by this organism for feeding, movement, respiration and reproduction occur inside the coelenteron. This body cavity functions in circulation, digestion, excretion and hydrostatic skeletal support, to list a few things. More detail for each mechanism can be found in the links below.
Few behavioural observations were made, mainly due to inactivity of the animal during study.
Feeding
With an oral-aboral axis and radial symmetry, these organisms have a 360° view of the environment around them. By using the tentacles that surround their oral disc, they are able to opportunistically capture prey in the water surrounding them. When a tentacle senses a prey item, the cnidocyst cells are fired, and the tentacles move the captured prey item to the mouth, where it is consumed (Mather & Bennett 1994; Ruppert et al. 2004). After digestion has taken place, the waste product (ammonia) is expelled from the polyp by diffusion through the body wall (Ruppert et al. 2004).
Protopalythoa species are polytrophic, which means that they use multiple methods to gain nutrients required for survival; they are not only micro-predators, as they are also able to gain energy from the process of photosynthesis, due to symbiotic zooxanthellae (Gosliner et al. 1996).
This particular species of Protopalythoa had evidence of chlorophyll in its body, which illustrates that energy would have been provided for this organism from photosynthetic processes—for more information, visit the 'Ecology' page.
Protopalythoa species utilize two main forms of reproduction: sexual and asexual. The process of budding or fragmentation is common in all polyps (Mather & Bennett 1994; Ruppert et al. 2004; Ryland 1997), with budding referring to the polyp creating a genetically identical copy of itself. Fragmentation, on the other hand, is the re-creation of the colony after damage has occurred, such as a piece of the substrate that the colony is occupying breaking off (Ruppert et al. 2004; Ryland 1997). This appears to be the most common method of zoanthid species spreading throughout a reef environment (Ryland 1997).
Sexual reproduction in this genus is carried out within the hermaphroditic colony (Ryland 1997), and results in the creation of a planula ‘zoanthella’ larva. A study by Babcock and Ryland (1990) demonstrated that after approximately 18 days, in suitable conditions these larvae would settle and metamorphose into adult forms.
Ryland (1997) concluded that in the Great Barrier Reef, Protopalythoa species and other zoanthid species spawn in the same pattern as other coral species—because of this, it has been suggested that many corals may be hybrids (Reimer et al. 2007). Due to continuously changing oceanic currents and other factors that would affect distribution of larvae as well as sperm and eggs, there is the potential for repeated sexual isolation and genetic differentiation (Reimer et al. 2007), which could help explain the plethora of morphotypes in Palythoa and Protopalythoa species.
Movement
The classic Cnidarian life history contains two main stages—a polyp or sessile stage, and a medusa or free-swimming stage (Gosliner et al. 1996). Because the class Anthozoa does not contain a medusoid stage in its life history (Gosliner et al. 1996; Mather & Bennett 1994), this Protopalythoa species remains in polyp form for its entire life history, with the exception of larval dispersal stages. As a result, this species does not move in the sense that it would decide to move to a different habitat. However, local movements in individuals of this species are made possible by the hydraulic pressure created inside the body (Erhardt & Knop 2005; Ruppert et al. 2004). Using this hydrostatic pressure, coupled with muscle fibres, polyps in Protopalythoa colonies are able to extend and retract their body and tentacles. Movement is only possible when the mouth is closed—otherwise the water that is put under pressure will rush out of the mouth during movement (Erhardt & Knop 2005; Ruppert et al. 2004). This is why the mouth opening, and the pharynx inside, is laterally compressed (or “flat”).

|
| The laterally compressed mouth opening is visible in this photograph, with the oral opening being a small slit. This prevents water from escaping when high pressures are created when the animal is moving its tentacles and body. |
This allows high pressure to be generated within the organism during movement, as the mouth is pressed closed by the pressure created, rather than being forced open, as would occur if the oral opening were a round shape (Erhardt & Knop 2005).
Respiration
Respiration in this species is carried out via diffusion, either over the body wall surface, or internally (Ruppert et al. 2004). Water flow into the coelenteron for respiration is driven by cilia that the line the siphonoglyph on the pharynx (Erhardt & Knop 2005). The continuous beating of the cilia causes a water current to form, helping to bring oxygen-rich water inside the animal.
Defences
Due to the fact that this Protopalythoa species doesn’t excrete a calcium skeleton (Davey 1998; Erhardt & Knop 2005; Mather & Bennett 1994), the ability for mechanical protection of the polyps in the colony is limited. Defence, from both predators and interspecies aggression, is therefore primarily achieved through the use of the phylum-specific sensory-effector cells called nematocysts (Reft & Daly 2012). Nematocysts are a defensive cell type in the Cnidarians, which occur in the body wall. In Protopalythoa species, these cells are present in the tentacles, and aid in prey capture and defence from predators and interspecies aggression (Reft & Daly 2012; Ruppert et al. 2004). A nematocyst is a cell consisting of a capsule with a lid (the apex) that has a tubule inside—this tubule is the ‘harpoon’ that is discharged when the organism senses a predator or prey item.
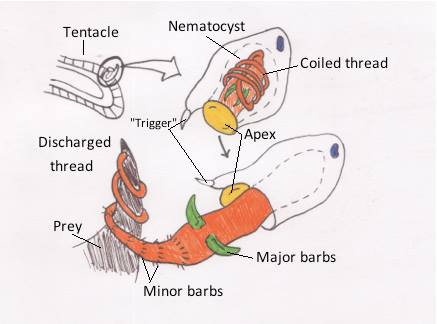
|
| Diagram illustrating a nematocyst cell before and after discharge. Adapted from Campbell et al. (2009). |
The apex of the nematocyst is either a hinged cap or three flaps that flex outward (Reft & Daly 2012). These cells are very energetically expensive for the animal, as each nematocyst can only be used once (Reft & Daly 2012; Ruppert et al. 2004).
In their study, Reft & Daly (2012) suggest that the structure of the nematocysts in Cnidarians could provide information important to classifying species, as there is a great amount of morphological variation (such as size, shape, and arrangement of spines) in nematocysts between orders in the phylum Cnidaria.
|